Xi Liu,
Lei Chen,
Yang Liu,
Tao Zhang ![]()
For correspondence:- Tao Zhang Email: zhangtaoasd01@126.com
Received: 11 February 2016 Accepted: 5 December 2016 Published: 31 January 2017
Citation: Liu X, Chen L, Liu Y, Zhang T. Tangeretin sensitises human lung cancer cells to TRAIL-induced apoptosis via ROS-JNK/ERK-CHOP pathway-mediated up-regulation of death receptor 5. Trop J Pharm Res 2017; 16(1):17-29 doi: 10.4314/tjpr.v16i1.4
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the efficacy of tangeretin, a bioactive flavonoid, as an enhancer of tumour necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis in human lung cancer cells (H1299 and H1975).
Methods: H1299 and H1975 cells were exposed to tangeretin (25, 50, or 100 µM), and its effect on cell viability was assessed by MTT assay. The cytotoxic effects of tangeretin in combination with TRAIL (25 ng/mL) were determined by live/dead assay and annexin V staining. Intracellular levels of reactive oxygen species (ROS) were determined by DHFDA-flow cytometry analysis. Western blotting was performed to assess the ex
Results: MTT assay revealed that tangeretin exerted potent cytotoxic effects and, in combination with TRAIL, exhibited striking apoptosis. Down-regulation of cell survival proteins (Bcl-2, Bcl-xL, survivin, xIAP, c-IAP-1, and c-IAP-2) and elevated ex
Conclusions: Tangeretin induces death receptors and enhances TRAIL-induced apoptosis through up-regulation of the ROS-JNK/ERK-CHOP pathways.
Introduction
Despite immense progression in the field of medicine, cancer remains a major threat to human health. Lung cancer is a prominent cause of cancer-related death worldwide [1]. Non-small cell lung cancer (NSCLC), the primary type of lung cancer, accounts for approximately 85 % of all cases, and small cell lung cancer (SCLC) accounts for 15 % [2]. Despite advancements in chemotherapy and radiation therapy, the long-term survival of patients with NSCLC remains low, with a 5-year survival rate of about 15 % [3]. Induction of apoptosis is crucial in cancer therapy. Much research has recently focused on strategies that specifically target molecules directly involved in the apoptotic pathways, thereby enhancing cancer cell death.
Tumour necrosis factor-related apoptosis-inducing ligand (TRAIL or Apo2 ligand) is a member of the tumour necrosis factor superfamily. TRAIL selectively induces apoptosis of cancer cells with negligible effects on normal cells [4,5]. TRAIL exerts its effects through interaction of its ligands with two membrane receptors, TRAIL-R1 (DR4) and TRAIL-R2 (DR5) [6]; the antagonistic transmembrane decoy receptor (DcR)-1 and DcR2 [7]; and the soluble receptor osteoprotegerin, which lacks the transmembrane domain [8].
The interaction of TRAIL with DR4 and DR5 leads to an association with Fas-associated death domain and caspase-8, forming a death-inducing signal complex that results in activation of an initiator caspase (caspase-8) and effector caspases (caspases-3, -6, -7, and -9), thereby inducing apoptosis [9,10]. TRAIL-induced preferential apoptosis of cancer cells makes this agent crucial in cancer treatment. However, many cancer cells acquire resistance to TRAIL by multiple mechanisms [11]. The mechanisms underlying TRAIL resistance include down-regulation of DRs and up-regulation of decoy receptors [11,12] and various anti-apoptotic proteins, including cellular FLICE-like inhibitory protein (cFLIP(L)), which competes with caspase-8 [13] and inhibitor of apoptosis proteins (IAPs) [14]. Thus, identification of TRAIL sensitizers that will overcome TRAIL resistance is a valuable approach in cancer therapy.
Several studies have demonstrated that plant-derived compounds effectively sensitise cancer cells to TRAIL-induced apoptosis [15,16]. Tangeretin (4′,5,6,7,8-pentamethoxyflavone), widely present in citrus fruits, possesses numerous bioactive properties, such as anti-inflammatory [17], anti-proliferative [18], and neuroprotective [19] activities. In the present study, we investigated whether tangeretin effectively sensitises and enhances human lung cancer cells to TRAIL-induced apoptosis.
Methods
Cell lines
The human lung cancer cells H1299 and H1975 were obtained from ATCC and maintained according to the manufacturer’s instructions. In brief, the cells were cultured in RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% foetal bovine serum, 100 units/mL of penicillin, and 100 µg/mL of streptomycin.
Chemicals and reagents
Tangeretin (Sigma-Aldrich, St. Louis, Mo, USA) and human trail (Peprotech, Rocky Hill, NJ, USA) were used. A cell apoptosis elisa detection kit was obtained from Roche. 2’,7’-dichlorofluorescein diacetate (dcf-da) (Molecular Probes Inc., Eugene, OR, USA) was used in the study. Antibodies to Bcl-xL, Bcl-2, survivin, Bax, cFLIP, CEBP homologous protein (CHOP), xIAP, cIAP-1, cIAP-2, caspase-3, caspase-8, caspase-9, DcR1, DcR2, DR4, DR5 (Cell Signaling Technology, Danvers, MA, USA), JNK, phospho-JNK, ERK1/2, phospho-ERK1/2, p38, phospho-p38, and β-actin (Santa Cruz Biotechnology, Santa Cruz, CA,USA) were used for expression analysis. Other chemicals and reagents used in the study were of analytical grade and were purchased from Sigma-Aldrich unless otherwise mentioned.
Cell viability assay
The cytotoxic effects of tangeretin (25–100 µM) on human lung cancer cells (H1299 and H1975) were assessed by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. In brief, the cells were seeded into 96-well plates (0.5 × 106cells/well) and incubated for 24 h followed by exposure to various concentrations of tangeretin for 24 h. After incubation, the cells were treated with MTT. After 4 h, the formazan crystals were dissolved in DMSO and the absorbance was measured at 570 nm using a Multiskan Spectrum (Thermo Electron Co., Vantaa, Finland).
Live/dead assay
A live/dead assay was performed to further assess apoptosis. The experiment was carried out using a Live/Dead assay kit (Invitrogen). The assay employs calcein-AM, a non-fluorescent polyanionic dye that determines the membrane integrity as a measure of cell viability. In brief, the cells with tangeretin (25, 50 or 100 µM) and/or TRAIL (25 ng/mL) were stained with Live/Dead reagent (5 μmol/L ethidiumhomodimer and 5 μmol/L calcein-AM). The cells were then incubated at 37 °C for 30 min. Apoptosis was assessed by measuring the fluorescence under a fluorescence microscope (Labophot-2; Nikon, Tokyo, Japan). Live cells retained the calcein-AM dye and produced intense green fluorescence, while the ethidiumhomodimerbecame bound to nucleic acids inside the dead cells and produced bright red fluorescence.
Analysis of apoptosis by Annexin V assay
Human lung carcinoma cells were incubated with tangeretin (50 or 100 µM) and/or TRAIL (25 ng/mL) for 24 h, and apoptosis was detected using an Annexin V-FITC detection kit II (BD Biosciences Pharmingen, San Diego, CA, USA). After incubation, the cells were treated with 5 μL annexin V-FITC reagent, incubated for 30 min at room temperature, and analysed using a flow cytometer (FACS Calibur; BD Biosciences).
Analysis of DR4 and DR5 expression
The influence of tangeretin (25, 50, and 100 µM) on the cell-surface expression of death receptors (DR4 and DR5) in the lung cancer cells was assessed using mouse anti-human DR4 or DR5 monoclonal antibodies conjugated with phycoerythrin (R&D Systems,Minneapolis, MN, USA). Cells treated with tangeretin for 24 h were incubated with antibodies for 45 min at 4 °C. The expression was analysed by flow cytometry [20].
RT-PCR analysis
The mRNA expression of DR5 and CHOP was assessed by performing RT-PCR. Total RNA was extracted from the cells after exposure to tangeretin (25, 50, and 100 µM) for 24 h using TRIzol reagent (Invitrogen). The total RNA was amplified by PCR [21] using the following primers: DR5, sense 5’-AAGACCCTTGTG CTCGTTGTC-3’ and anti-sense 5’-GACACATT CGATGTCACTCCA-3’; CHOP, sense 5’-CAACTGCAGAGATGGCAGC TGA-3’ and antisense 5’-CTGATGCTCCCAATT GTTCAT-3’; and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), sense 5’-GTCTTCACCACCATGGAG-3’ and antisense 5’-CCACCCTGTTGCTGTAGC-3’. The relative expression levels of CHOP and DR5 genes were normalised with the expression of GAPDH.
Transfection with siRNA
The influence of tangeretin on DR5 and CHOP expression following silencing with respective siRNA (Santa Cruz Biotechnology) was assessed. The H1299 and H1975 cells were transfected with siRNA oligonucleotides (30 nmol/L) using lipofectamine 2000 (Invitrogen) according to the manufacturer’s instructions. Following transfection, the cells were treated with tangeretin (100 µM) for 12 h and incubated with TRAIL (25 ng/mL) for 24 h [22]. After these treatments, the cells were collected and analysed for expression using western blotting, and apoptosis was assessed by the Live/Dead assay.
Measurement of reactive oxygen species
To detect intracellular reactive oxygen species (ROS) in the lung cancer cells, the cells were treated with 20 µM DCF-DA for 15 min at 37°C. Following DCF-DA treatment, the cells were incubated with various concentrations of tangeretin (25, 50, or 100 µM) for 30 min. The increase in fluorescence resulting from the oxidation of DCF-DA to DCF was analysed by flow cytometry at 530 nm as previously described [23]. Data were analysed for at least 10,000 cells at a flow rate of 250 to 300 cells/s.
Western blotting
The cells treated with tangeretin (25, 50, or 100 µM) and/or TRAIL (25 ng/mL) for 24 h were subjected to western blot analysis to assess the expression of proteins. Western blot analysis was carried out as previously described by Yang et al [24]. Following treatment with tangeretin and TRAIL, the cells were incubated in 0.5 mL of ice-cold whole-cell lysate buffer (5 M NaCl, 10 % Nonidet P-40, 0.2 M sodium orthovanadate, 0.1 M EGTA, 0.5 M EDTA, 0.1 M phenylmethyl-sulfonyl fluoride, 1 M sodium fluoride, 1 M HEPES 2 µg/mL aprotinin, and 2 µg/mL leupeptin) on ice for 30 min. The isolated protein concentrations were determined using a Bio-Rad assay kit (Bio-Rad, Hercules, CA, USA). Equal amounts (60 µg) of isolated proteins were fractionated using SDS-PAGE and transferred onto polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA), following incubation with respective antibodies. The immunoreactive bands were detected and analysed using an ECL Advance western blot analysis system (Amersham Pharmacia Biotech Inc., Piscataway, NJ, USA). The band densities were normalised to those of control β-actin using anti-β-actin antibody (Cell Signaling Technology).
Statistical analysis
SPSS software (ver. 22.0; IBM Corp., Armonk, NY, USA) was used for analysis. The experimental data are given as means ± SD (n = 3 or 6). The means of the various groups were compared by one-way ANOVA followed by Duncan’s multiple range test (DMRT) as a post-hoc analysis. Differences with a p value of < 0.05 were considered statistically significant.
Results
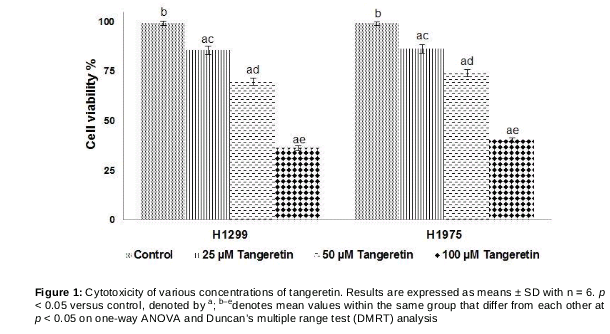
Tangeretin inhibited cancer cell proliferation
Tangeretin at 25, 50, and 100 µM significantly (p < 0.05) inhibited the proliferation of H1975 and H1299 cells (). The cell viability percentage decreased in a concentration-dependent manner. The highest dose (100 µM) exhibited maximum inhibitory effects as compared with the lower doses (25 and 50 µM).
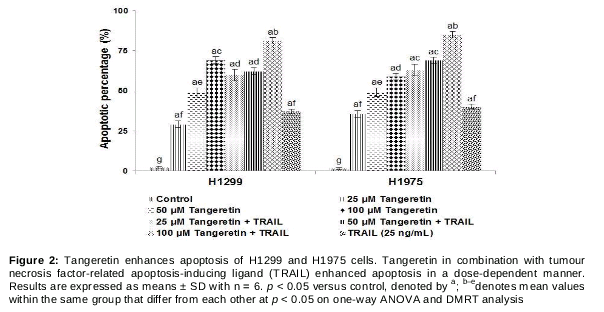
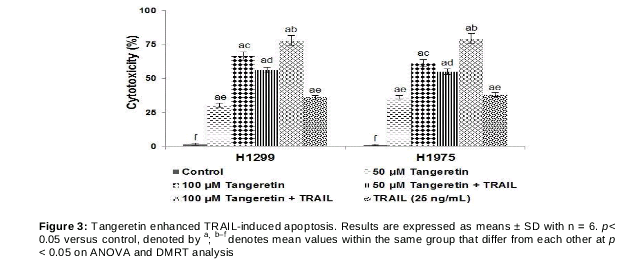
Tangeretin enhances TRAIL-induced apoptosis of lung cancer cells
We also examined whether tangeretin was able to enhance and sensitise lung cancer cells to TRAIL-induced apoptosis by the Live/Dead assay and Annexin V staining. The Live/Dead assay was performed after exposure to 25, 50 or 100 µM tangeretin. Exposure to the 100-µM concentration resulted in a drastic increase in apoptosis (69.09 % of H1299 cells and 58.93 % of H1975 cells) (). Tangeretin at the 100-µM concentration induced higher apoptotic cell counts than did lower concentrations. Interestingly, however, combined exposure to TRAIL and tangeretin at 100 µM resulted in a multi-fold increase in cytotoxicity. The apoptosis percentage increased to 81.19 % in H1299 cells and 84.90 % in H1975 cells (), while TRAIL alone induced apoptosis of about 37.18 and 40.24 % in H1299 and H1975 cells, respectively.
We further analysed the efficacy of tangeretin in enhancing TRAIL-induced apoptosis by Annexin V staining assay. The assay examines the integrity of the cell membrane as a measure of cell viability. The translocation of phosphatidylserine from the cytoplasmic interface to the extracellular surface of the membrane is a preliminary indicator of apoptosis. The results presented in show that tangeretin was able to markedly (p < 0.05) increase the apoptosis percentage with combined exposure to TRAIL and tangeretin at 50 and 100µM compared with exposure to either TRAIL or tangeretin. These observations suggest that tangeretin at the tested doses significantly enhanced sensitivity to TRAIL and improved TRAIL-induced apoptosis.
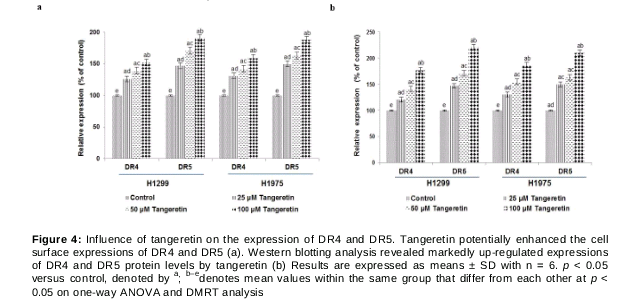
Tangeretin potentially up-regulates the expression of DR4 and DR5
Studies have shown that down-regulation or loss of DR4 and DR5 expression is a major mechanism involved in TRAIL resistance [10,11]. We observed significantly (p < 0.05) enhanced cell surface expression of DR4 and DR5 upon treatment with tangeretin in a dose-dependent manner (a). Furthermore, expression at the protein level was detected by western blot analysis. The results revealed similarly increased expression of DR4 and DR5 upon exposure to tangeretin (b), suggesting the efficacy of tangeretin in up-regulating the expression of DR4 and DR5, thereby increasing apoptosis. Additionally, 100 µM of tangeretin increased the expression when compared with lower doses.
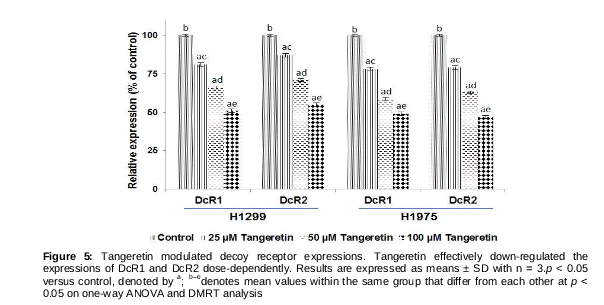
Influence of tangeretin on the expression of decoy receptors
The enhanced expression of antagonistic decoy receptors of TRAIL (DcR1 and DcR2) is reportedly involved in TRAIL resistance [12]. We observed that tangeretin, at all tested doses, was able to potentially supress the expression of DcR1 and DcR2.
However, the effects were more pronounced on DcR1 than DcR2, and the 100-µM dose exhibited maximal effects (). Thus, by down-regulating DcR1 and DcR2 and inducing DR4/5 expression, tangeretin effectively aids in activating apoptosis of lung cancer cells.
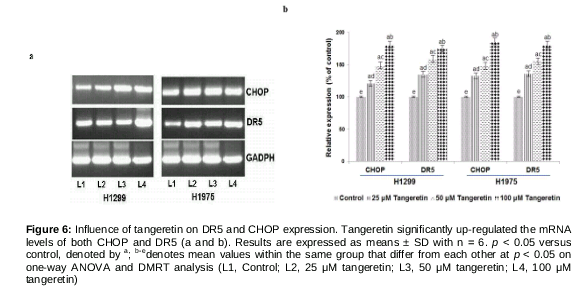
CHOP mediates tangeretin-induced up-regulation of death receptors
It has been demonstrated that the induction of DRs by various stimulants is mediated through the activation of CHOP [25,26]. We investigated whether tangeretin influenced CHOP expression and whether CHOP is also involved in DR5 expression and tangeretin-induced apoptosis. RT-PCR analysis revealed enhanced DR5 and CHOP mRNA levels (a and b). In line with the CHOP mRNA levels, western blot analysis also revealed significantly (p < 0.05) enhanced CHOP protein expression. These observations indicate a positive influence of tangeretin on CHOP expression.
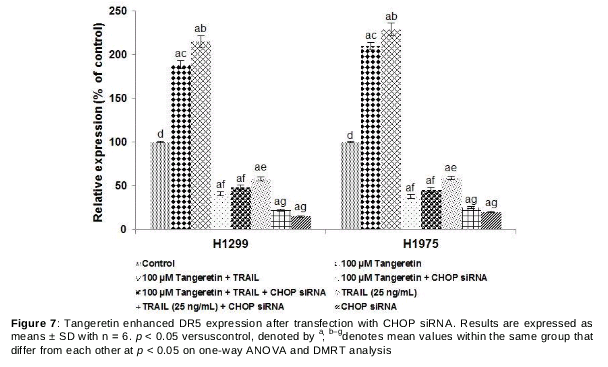
siRNA was used to further confirm whether tangeretin was able to mediate DR5 expression via CHOP and to determine whether CHOP-DR5 is involved in tangeretin-mediated apoptosis. Interestingly, DR5 expression decreased upon transfection with CHOP siRNA (), and this decreased expression markedly affected the apoptosis percentage (Figs. 8 and 9). Significantly reduced apoptosis of lung cancer cells was observed on transfection with both CHOP siRNA and DR5 siRNA. However, tangeretin treatment considerably enhanced DR5 expression even in the presence of CHOP siRNA and enhanced the apoptosis percentage to some extent, while the percentage was lower than that in the absence of siRNA. These observations indicate that tangeretin-induced apoptosis occurs in part through direct stimulation of apoptosis and is in part mediated by CHOP. Additionally, CHOP is critical in DR5-mediated TRAIL-induced apoptosis.
Tangeretin modulates expression of various cell survival proteins
Numerous studies have demonstrated that over-expression of cell survival proteins, such as IAPs, survivin, cFLIP, Bcl-2, and Bcl-xL, is involved in the development of TRAIL resistance [27,28]. We observed significantly (p < 0.05) increased expression of cell survival proteins in H1299 and H1975 cells (0). However, tangeretin treatment at 25, 50, or 100 µM resulted in a decline in the expression levels in a dose-dependent manner.
Tangeretin up-regulates expression of pro-apoptotic proteins
We observed a remarkable increase (p < 0.05) in the expression of caspase-3, -8 and -9 in H1299 and H1975 cells on exposure to tangeretin (1); pro-apoptotic protein Bax was also up-regulated. The expression was dose-dependently regulated by tangeretin.
Tangeretin-induced up-regulation of TRAIL receptors is mediated by MAPKs
We investigated whether activation of MAPKs (JNK, ERK, and p38MAPK) is involved in tangeretin-induced DR5 induction. We observed a significant (p < 0.05) increase in the activation of ERK and JNK, as evidenced by raised phosphorylation levels. The 50- and 100-µM doses were more effective in activating JNK and ERK than was the 25-µM dose. Further, ERK levels were higher as against JNK. While the levels of p38 were noticeably increased by tangeretin, there was no significant increase, suggesting that the activation of JNK and ERK is involved more critically in tangeretin-mediated apoptosis than p38.
We also observed a drastic reduction in the expression of DR5 and DR4 in the presence of inhibitors of JNK (SP600125) and ERK1/2 (PD98059). However, the expression was not significantly affected by the presence of an inhibitor of p38 MAPK (SB202190). Tangeretin induced the expression of TRAIL receptors, even in the presence of inhibitors, in a dose-dependent manner. These observations illustrate that the activation of JNK and ERK is more closely involved in the up-regulated expression of DR4 and DR5.
Involvement of ROS in tangeretin-mediated TRAIL-induced apoptosis
ROS are reportedly involved in the induction of DRs [20]. We observed enhanced ROS levels upon exposure to tangeretin (3). The increase, however, was dose-dependent. These observations suggest the possible involvement of ROS in up-regulation of DRs and ERK/JNK MAPKs.
Discussion
Despite the promising effects of TRAIL in cancer cell therapy, many reports have described human tumours developing resistance to TRAIL [10,11] through various mechanisms. Altered expression of DRs and the Fas-associated death domain,as well as overexpression of anti-apoptotic proteins, such as Bcl-2, survivin, cFLIP [11,13,27,28], and xIAP, have been observed in many tumour cell lines exhibiting TRAIL resistance [28].
Thus, strategies to overcome this dysregulation and sensitise cells to TRAIL and possibly enhance its apoptotic effects are crucial and of tremendous value. Compounds of plant origin are known to effectively sensitise cells to TRAIL-induced cancer cell cytotoxicity [15,20]. Here, we investigated whether tangeretin was able to sensitise and enhance TRAIL-induced apoptosis in the human lung cancer cells H1299 and H1975.
Treatment with tangeretin at 25 to 100 µM reduced the viability of H1299 and H1975 cells, indicating its potent anti-proliferative efficacy. The combined exposure to TRAIL and tangeretin dramatically increased the apoptotic cell counts compared with treatment with tangeretin or TRAIL separately. This suggests that tangeretin was able to effectively enhance the effects of TRAIL; in addition, tangeretin down-regulated the cell survival proteins Bcl-2, Bcl-xL, xIAP, cFLIP, cIAP-1, cIAP-2, and survivin. These proteins have been reported to be associated with TRAIL resistance [27,28]. Thus, decreased expression of anti-apoptotic proteins aids in TRAIL-induced apoptosis. Previous studies have demonstrated that down-regulation of survivin, Bcl-xL, and Bcl-2 promotes sensitivity to TRAIL [27,28].
xIAP contributes to TRAILresistance by inhibiting caspase-3, -7, and -9 [28], and cFLIP inhibits caspase activation by competing for the Fas-associated death domain [13]. Thus, effective suppression of cFLIP and IAPs by tangeretin could potentially contribute to sensitising cancer cells to TRAIL. In the present study, elevated levels of caspase-3, -8 and -9 were observed upon tangeretin exposure. This up-regulated expression could be due to the inhibition of xIAP and cFLIP. Increased expression of the pro-apoptotic protein Bax also aids in enhancing TRAIL-mediated apoptosis. The modulations observed in the expression patterns were dose-dependent, with 100-µM of tangeretin exhibiting maximal effects; further combination with TRAIL showed an additional influence. Phytochemicals, such asguggulsterone and nimbolide [15,25], enhanced TRAIL-mediated cell death via up-regulation of caspases and suppression of cell survival proteins.
Binding of TRAIL with the death receptors DR4 and DR5 triggers TRAIL-induced apoptotic signalling, leading to the activation of executor caspase. Dysregulated expression levels have been reported in several TRAIL-resistant cancers [29], and subsequent up-regulation of the receptors promotes apoptosis [11,25]. Significantly up-regulated cell surface expression of DR4 and DR5 was observed upon tangeretin treatment. While tangeretin exposure caused elevated expression of DR4 and DR5, the expression of TRAIL decoy receptors DcR1 and DcR2 was dose-dependently reduced at the protein level. Mellier et al [30] showed that sensitivity to TRAIL-mediated apoptosis is influenced by the over-expression of TRAIL-decoy receptors. The down-regulated expression of DcR1 and DcR2, along with elevated DR4 and DR5 expression upon exposure to tangeretin, effectively contributes to enhance TRAIL-mediated apoptosis.
Furthermore, DR5 expression was up-regulated by tangeretin even at the gene level, as evidenced by increased DR5mRNA levels. The mRNA levels of CHOP were also enhanced upon tangeretin exposure. CHOP is a key transcriptional factor that regulates DR5. CHOP binds to the DR5 promoter region and up-regulates DR5 expression [17]. The enhanced CHOP levels on tangeretin exposure suggest that tangeretin modulates DR5 expression via CHOP. This was further confirmed when CHOP expression was silenced using CHOP siRNA, which resulted in significant inhibition of DR5 expression; however, tangeretin was able to considerably enhance DR5 expression even with CHOP siRNA. Silencing of DR5 and CHOP genes resulted in a sharp decline in apoptosis levels, reflecting the vital roles of DR5 and CHOP in TRAIL-mediated cell death. To an extent, tangeretin exposure was able to increase the apoptosis levels even in the presence of DR5 siRNA and CHOP siRNA. These observations suggest that tangeretin was able to potentiate the apoptosis-mediating effects of TRAIL by modulating DR5 and CHOP.
We also observed that tangeretin was able to induce ROS production in H1299 and H1975 cells. The observed ROS levels could have also induced CHOP expression, because CHOP is a major stress-regulated protein [25]. Several studies have reported that chemotherapeutic agents enhance DR5 expression and TRAIL-mediated apoptosis via ROS-dependent pathways [15,25,26].
ROS reportedly trigger various signal transduction pathways that are involved in cell growth, differentiation, or death. MAPKs are important downstream mediators of ROS-induced signalling [26]. ERK, JNK, and p38 MAPK have been found to be activated in TRAIL-mediated apoptosis [15,30] and are involved in TRAIL receptor induction [15,26]. In our study, the significant increases in the activation of JNK and ERK, along with a considerable increase in p38 MAPK activation on exposure to tangeretin, could have been either due to direct stimulation of tangeretin and/or in part indirectly due to ROS. We also observed suppression of DR4 and DR5 in the presence MAPK inhibitors, which was noticeably enhanced by tangeretin.
Conclusion
Tangeretin up-regulates death receptors and enhances TRAIL-mediated apoptosis via ROS-CHOP signalling and MAPKs. Thus, tangeretin should be further explored as a potent candidate for combined therapy with TRAIL for induction of apoptosis of cancer cells.
Declarations
Acknowledgement
References
Archives


News Updates